|
ГЕОЛОГИЧЕСКИЙ ИНСТИТУТ На правах рукописи
ЭОПЛЕЙСТОЦЕНА ЮГА ВОСТОЧНОЙ ЕВРОПЫ ( по мелким млекопитающим )
диссертации на соискание ученой степени кандидата геолого-минералогических наук
Научный руководитель
|
|
Работа выполнена в Геологическом институте РАН
на заседании диссертационного совета Д.002.215.03 по
палеонтологии и стратиграфии в Геологическом институте РАН
С диссертацией можно ознакомиться в отделении геологогической литературы Библиотеки по естественным наукам РАН (Старомонетный пер., д. 35)
Автореферат разослан 10 января 2002 года.
|
|
Актуальность темы. В настоящее время одной из главнейших задач стратиграфии континентальных отложений позднего кайнозоя является создание общей детальной биостратиграфической шкалы Евразии. Большая скорость эволюции, быстрота расселения млекопитающих выдвигают их на ведущую роль в палеонтологическом обосновании стратиграфии. Из всех групп для этой цели особенно важны мелкие млекопитающие, поскольку их местонахождения встречаются чаще, чем крупных животных, они нередко бывают многослойными и доставляют серийный материал, повышающий надёжность биостратиграфических построений. Однако создание общей шкалы осложняется палеозоогеографическими различиями отдельных регионов. Поэтому для построения такой шкалы необходимым этапом служит разработка региональных шкал и последующая их корреляция между собой. Актуальность настоящей работы определяется попыткой разработки одной из таких региональных шкал, охватывающей интервал времени примерно от 3,5 до 0,8 млн. лет – средний плиоцен - эоплейстоцен. Цели и задачи исследований. Основная цель исследований – создание зональной шкалы по мелкими млекопитающим для континентальных отложений среднего плиоцена – эоплейстоцена юга Восточной Европы. Для построения шкалы были выбраны полёвки (Arvicolinae), исходя из того, что особенностью фауны мелких млекопитающих умеренных широт Евразии в среднем плиоцене – эоплейстоцене является повсеместное распространение корнезубых полёвок. В соответствии с поставленной целью решались следующие задачи: 1. Анализ геологического строения основных опорных местонахождений юга Восточной Европы и таксономического состава сообществ мелких млекопитающих. 2. Ревизия фаунистического материала из некоторых опорных местонахождений европейской части России, Украины и Молдавии, собранного другими исследователями. 3. Выяснение стратиграфического распространения руководящих форм полёвок и установление последовательности фаун и местонахождений. 4. Разработка методики для построения зональной шкалы. 5. Обоснование возраста выделенных подразделений и их корреляция с различными стратиграфическими шкалами. 6. Описание палеонтологического материала по основным группам полёвок. Материал. Автором собран палеонтологический материал из семи местонахождений Северного Причерноморья (Волна, Крыжановка, Хаджибейский лиман), Приазовья (Цимбал, Тиздар, Ахтанизовская), Северного Кавказа (Псекупс) и изучена геология этих разрезов. Кроме того Л.П.Александрова любезно передала автору для обработки коллекции ископаемых грызунов из местонахождений Молдавии (Рипа Скорцельская, Долинское, Этулия), Приазовья (Ливенцовка), Северного Кавказа и Закавказья (Сабля, Псекупс, Кушкуна), Северного Причерноморья (Веселовка). В распоряжении автора были также коллекции из сборов В.А.Присяжнюка и А.А.Круковера из местонахождений Широкино и Ливенцовка. Все перечисленные коллекции хранятся в ГИН РАН. Благодаря любезности В.Л.Яхимович и В.П.Сухова автор имел возможность ознакомиться с коллекциями из местонахождений Башкирского Предуралья (Аккулаево и Симбугино), хранящимися в Институте геологии Башкирского НЦ УрО РАН (г. Уфа). Для сравнительных целей в работе привлекался материал, просмотренный автором из ряда других местонахождений: Котловина, Черевичное, Жевахова гора (Украина), Нагавская и др. (бассейн реки Дон), Тегелен (Нидерланды), Фрехен и Хамбах (Германия) и ряда других. Всего изучено около 6 тысяч ископаемых остатков, принадлежащих 95 формам мелких млекопитающих, из них 45 полёвок. Проведено свыше 15 тысяч измерений элементов зубов. Защищаемые положения. 1. Зональная биостратиграфическая шкала континентальных отложений среднего плиоцена – эоплейстоцена юга Восточной Европы, разработанная на основе эволюционных изменений в четырёх филетических линиях полёвок (Borsоdia, крупные Mimomys, мелкие Mimomys и Pitymimomys) с учётом количественных оценок морфологических признаков (главным образом, степени гипсодонтии). 2. Корреляция выделенных зон с подразделениями стратиграфических шкал Восточного Паратетиса, континентальных отложений Западной Европы и магнитохронологической шкалы. Сопоставление опорных местонахождений фауны Восточной Европы с главнейшими местонахождениями Центральной и Северо-Западной Европы. 3. Количественные критерии определения переотложенных форм полёвок в местонахождениях аллювиального типа. 4. Монографическое описание 26 видов и подвидов полевок, принадлежащих к 7 родам и подродам, в том числе одного нового рода и одного нового вида. Научная новизна. Впервые для построения зональной биостратиграфической шкалы привлечены количественные оценки морфологических признаков на зубах полёвок, определяющие степень эволюционной продвинутости форм. Статистический анализ изменчивости морфологических признаков позволил предложить количественные критерии определения переотложенных форм полёвок в «смешанных» фаунах. На юге Восточной Европы установлено более раннее появление родов Allophaiomys и Prolagurus, чем было принято до настоящего времени. В подсемействе Arvicolinae выделены новые род и вид. Практическая значимость работы. Предложенная зональная шкала может быть использована для палеонтологического обоснования стратиграфии континентальных отложений среднего плиоцена - эоплейстоцена Восточной Европы. Ревизия фаун полёвок из ряда опорных местонахождений позволяет внести коррективы в некоторые существующие стратиграфические схемы. Апробация работы. Основные результаты исследований освещены в 26 статьях и 13 тезисах докладов и докладывались на конкурсных сессиях Геологического института РАН, на заседаниях Неогеновой и Четвертичной комиссий МСК, Московского общества испытателей природы, семинаре по мелким млекопитающим РМСК по Центру и югу Русской платформы, на Всесоюзном совещании по изучению четвертичного периода (Таллин, 1990), Всероссийских совещаниях по изучению четвертичного периода (Москва, 1994, Санкт-Петербург, 1998), на Международной конференции «Неогеновые и четвертичные млекопитающие Европы» (Краков, 1994), Международном симпозиуме «The Dawn of the Quaternary» (Нидерланды, 1996), заседании Международной рабочей группы «Происхождение, эволюция, систематика, стратиграфическое и географическое распространение Allophaiomys (Rodentia, Arvicolinae)» (Москва, 1997). Структура работы. Работа состоит из введения, трех глав, заключения, списка использованной литературы и приложения. Общий объем работы страниц, включая страниц текста, рисунков, таблиц. Список литературы включает названий. Приложение (описание палеонтологического материала) содержит страниц текста, включая 134 таблицы и 44 рисунка. Работа выполнена в лаборатории стратиграфии четвертичного периода Геологического института РАН под руководством д.г.-м..н. Э.А.Вангенгейм, которой автор выражает искреннюю благодарность. В палеонтологических исследованиях автор получал консультации к.г.-м.н. В.С.Зажигина, без которых работа была бы невозможной. Автор весьма признателен также д.г.-м..н. М.А.Певзнеру за всестороннюю помощь и поддержку в ходе работы. Автор благодарен д.г.-м..н. Е.В.Девяткину за создание благоприятных условий при написании работы, коллегам по лаборатории за ценные замечания и другую помощь в процессе подготовки диссертации; д.б.н. А.О.Аверьянову, д.б.н. А.К.Агаджаняну, к.г.-м.н. Л.П.Александровой, д.г.-м.н. Л.И.Алексеевой, к.б.н. В.С.Байгушевой, к.б.н. А.В.Бородину, д.б.н. И.М.Громову, к.б.н. В.И.Жегалло, к.г.н. Н.Е.Казанцевой, к.г.-м.н. Т.В.Кузнецовой, д.г.н. А.К.Марковой, к.б.н. В.А.Несину, П.А.Никольскому, к.б.н. А.Ю.Пузаченко, д.б.н. Л.И.Рековцу, к.г.-м.н. В.П.Сухову, к.б.н. В.В.Титову, акад. НАНУ В.А.Топачевскому, к.б.н. Л.А.Тютьковой, к.г.-м.н. К.И.Шушпанову, к.б.н. А.Г.Яковлеву, д.г.-м.н. В.Л.Яхимович, профессору К. Ковальскому, доктору А.Надаховскому, профессору Р.Мартину, доктору А.ван дер Мелену, доктору Х. де Бруйну, доктору Т. ван Колфсхотену, проф. В.фон Кенигсвальду за предоставление как оригинального материала, так и сравнительных коллекций и обсуждение отдельных аспектов работы и другую помощь; чл.-корр. НАНУ В.Н.Семененко, к.г.-м.н. Е.В.Иосифовой, д.б.н. И.Г.Тактакишвили и д.г.н. А.Л.Чепалыге за определение остатков моллюсков; д.б.н. И.Я.Павлинову за помощь в работе с коллекциями по современным мелким млекопитющим; Т.А.Синяковой за большую техническую помощь. |
|
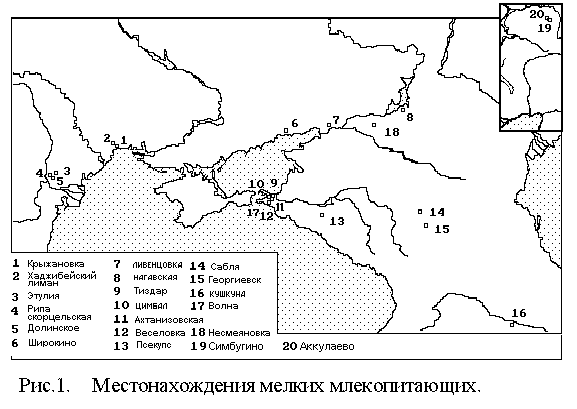
Глава 1. Геологическое строение опорных местонахождений фауны млекопитающих юга Восточной Европы. В работе использована стратиграфическая схема МСК (1995, 1998). Положение региоярусов Каспийского и Эвксинского бассейнов в магнитохронологической шкале принято по М.А.Певзнеру (1982, 1989). Корреляция описываемых разрезов и фаунистических подразделений проведена со схемами В.И. Громова (1948), М.Кретцоя (Kretzoi, 1991). А.Аццароли (Azzaroli et al., 1982; Sardella at al, 1998), П. Мэна (Mein, 1975) и О. Фейфара и др. (Feifar et al., 1998). В главе приведено геологическое описание разрезов, положение в них костеносных горизонтов и списки фауны мелких млекопитающих. Семь разрезов исследованы автором: Крыжановка, Хаджибейский лиман, Цимбал, Ахтанизовская, Тиздар, Волна, Псекупс. Геологическое описание 11-ти разрезов, фауну из которых изучал автор, приведено по литературным данным: Долинское, Рипа Скорцельская, Кушкуна, Этулия (Константинова, 1965; Путеводитель…, 1972, 1982), Симбугино, Аккулаево (Фауна и флора…, 1972, 1977) Широкино (Топачевский, 1965; Мацуй, Моськина, 1988), Сабля, Георгиевск (Лебедева, 1978), Веселовка (Александрова, 1975), Ливенцовка (Александрова, 1976) (Рис.1) . К среднему плиоцену относятся костеносные отложения разрезов Долинское, Рипа Скорцельская, Волна, Широкино, Кушкуна, Симбугино, Аккулаево (нижний горизонт) и Веселовка. Костеносные горизонты в разрезах Волна и Веселовка приурочены к солоноватоводным отложениям нижнего куяльника, а в разрезах Кушкуна и Аккулаево – к солоноватоводным отложениям среднего акчагыла. К верхнему плиоцену принадлежат костеносные отложения разрезов Сабля, Крыжановка, Этулия-3, Ливенцовка, Псекупс, Георгиевск, Тиздар. В разрезе Сабля остатки млекопитающих найдены вместе с раковинами акчагыльских моллюсков времени максимальной трансгрессии акчагыльского моря. В Крыжановке и Тиздаре костеносные горизонты связаны с солоноватоводными отложениями верхнего куяльника. К эоплейстоцену относятся костеносные отложения в разрезах Аккулаево (демский и давлекановский горизонты), Ахтанизовская, Цимбал и Хаджибейский лиман. |
|
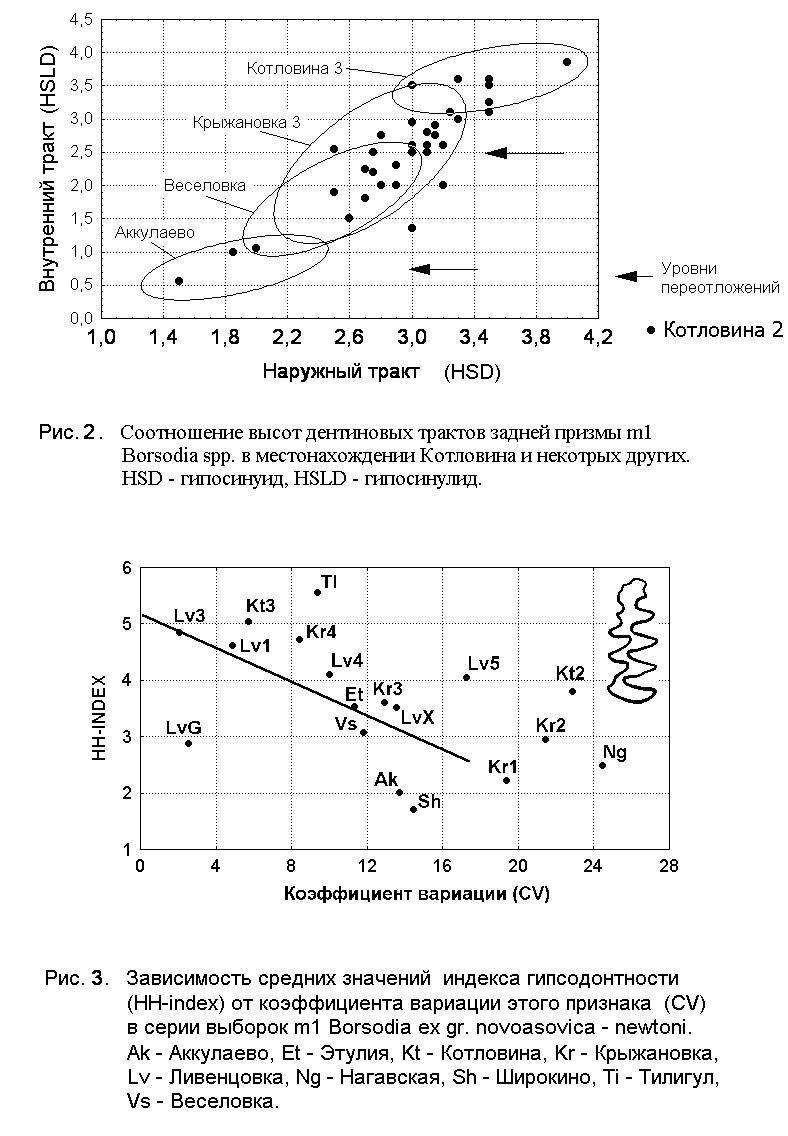
Глава 2. Методика исследований палеонтологического материала. Сбор и первоначальная обработка материала проводилась по традиционной методике. Терминология элементов зубов полевок, в том числе и генетическая терминология элементов зубов, приводится по системе А. ван дер Мелена (v.d.Meulen, 1973, 1974). Для характеристики стадии развития корней использованы определения Л. Мехели (Mehely, 1914), Н.Г. Смирнова и др. (1986), Н.Е. Казанцевой и автора (Kazantseva, Tesakov, 1998) с некоторыми изменениями. Терминология дентиновых трактов принята по Г.Рабедеру (Rabeder, 1981). В работе использована общепринятая система промеров зубов, а также некоторые промеры, предложенные А. ван дер Меленом (1973). В процессе работы было сделано свыше 15 тысяч промеров. Большинство из них перенесено в цифровые таблицы персонального компьютера. Обработка данных в цифровом формате велась с использованием программ MS Excel, Statgraphics и Statistica. Основной целью компьютерной обработки было вычисление статистических параметров варьирования данных (среднее, ошибка средней, стандартное отклонение, коэффициент вариации, индексы соотношений различных измерений), построение двухмерных графиков распределения измеряемых параметров, а также сравнение вариабельности признаков в различных выборках. В ряде случаев для анализа сходства ископаемых выборок применялся кластерный анализ. Гипсодонтия корнезубых полёвок. Эволюция зубной системы полёвок шла по пути формирования высококоронковых (гипсодонтных) призматических щечных зубов в условиях нарастающего остепнения умеренных широт Голарктики. В ряде эволюционных стволов полёвок этот процесс привёл к формированию полностью безкорнезубых моляров, растущих в течение всей жизни и никогда не образующих корни. Прогрессирующая гипсодонтия корнезубых полёвок может быть использована для целей детальной стратиграфии. В общем виде для членов одной эволюционной линии полёвок прослеживается закономерность – чем выше, гипсодонтнее коренные зубы, тем моложе геологический возраст вмещающих их отложений. Для оценки гипсодонтии полёвок используются различные методики: по соотношению высоты коронки и корней (Сухов, 1970; Топачевский и др., 1984), по сравнительному изучению стадий формирования корней коренных зубов современных и ископаемых полёвок (Kawamura, 1998; Kazantseva, Tesakov, 1998; Tesakov, 1998). На наш взгляд, наиболее перспективной оценкой гипсодонтии может служить высота дентиновых трактов, которая в различных модификациях используется многими исследователями (Hibbard, 1959; Hibbard, Zakrzhewsky, 1967; Nelson, Semken, 1970; Chaline, 1974; Агаджанян, 1976; van de Weerd, 1976; Зажигин, 1980 и др.). В настоящее время широко вошла в практику методика Г.Рабедера (Rabeder, 1981), который дал чёткую генетическую номенклатуру трактов, впервые использовал все коренные зубного ряда и предложил несколько удобных индексов. Так, для оценки гипсодонтии нижних моляров он ввёл НН-индекс – квадратный корень из суммы квадратов высот двух трактов задней призмы (гипосинуида и гипосинулида); для верхних коренных – PА-индекс, вычисляемый как квадратный корень из суммы квадратов высот трактов протокона и антерокона (протосинуса и антеросинуса). Г. Рабедер ввёл также синуграммы – двумерные диаграммы, в которых по оси абсцисс отложена величина индекса, а по оси ординат – длина коронки. В работе для оценки гипсодонтии используются как методы Г. Рабедера, так и диаграммы соотношения высоты различных трактов. В ряде случаев привлечение абсолютных величин дентиновых трактов, а не их индексных значений, позволяет получить более полную информацию о конкретной выборке. Регистрация переотложений. Известно, что аллювиальный тафономический тип захоронений особенно часто содержит материал, переотложенный из более древних слоёв в более молодые. Метод диагностики тафономических примесей для плио-плейстоценовых полёвковых фаун на основе изучения изменчивости дентиновых трактов был впервые применен В.С.Зажигиным для разделения фаунистических ассоциаций в сложнопостроенной аллювиальной толще в долине реки Битеке на юге Западной Сибири (Зыкин, Зажигин, Присяжнюк, 1987). Для плио-плейстоценовых полёвковых фаун юга Восточной Европы был проведен анализ изменчивости степени гипсодонтии коренных зубов. Детальная количественная оценка высоты дентиновых трактов зубов корнезубых полёвок показала неожиданно широкий разброс значений в некоторых выборках. Повышенная изменчивость может быть объяснена либо: 1) естественной широтой вариабельности признаков высоты трактов и/или 2) присутствием в захоронении нескольких родственных форм, различающихся по степени гипсодонтии, или 3) совмещением в захоронении путём переотложения нескольких эволюционных стадий одной филетической линии, которые различаются по степени гипсодонтии коренных. Специально проведенное автором изучение современных корнезубых полевок Clethrionomys glareolus показало, что разброс значений трактов невелик. Коэффициент вариации CV (отношение стандартного отклонения к средней, взятое в процентах) трактов современной выборки имеет значение 5 – 6 и никогда не достигает величин в 20 – 40, регистрируемых в некоторых ископаемых выборках. Средние же наблюдаемые значения CV для НН-индекса составляют для палеонтологического материала около 10 – 12. Версия о нескольких синхронных близкородственных формах, имеющих разную гипсодонтию, никогда полностью опровергнута быть не может. Однако наличие таких форм сразу в нескольких филетических линиях снижает её правдоподобность. В большинстве же случаев для аллювиальных захоронений верным оказывается третье предположение - о тафономическом совмещении остатков разных стадий одной филетической линии. Хорошим примером служит материал по полёвкам рода Borsodia из среднего слоя Котловины (коллекция Института зоологии НАН Украины) с CV-индексом 22,8. На рисунке 2 показано распределение трактов задней призмы m1 из этого местонахождения и наложены распределения высот дентиновых трактов из других местонахождений. По-видимому, в разбираемом случае можно говорить о двух уровнях переотложения: среднеплиоценового (соответствует распределению Аккулаево) и начала позднего плиоцена (соответствует распределению значений в местонахождении Крыжановка 3). Уровнем, на котором ископаемые остатки синхронны вмещающим отложениям, следует считать наиболее верхнюю часть распределения, близкую к ареалу трактов из верхнего слоя Котловины. О смешанном характере ассоциаций Котловины дополнительно свидетельствуют наличие раннеплиоценовых грызунов, таких как Trilophomys, и беспрецедентное разнообразие форм полевок Dolomys и Pliomys, по-видимому представляющих разновозрастные стадии развития единых филетических линий. Для фаун корнезубых полевок показателем переотложения может служить величина коэффициента вариации CV показателей гипсодонтии. Вероятно, критерий оценки эволюционной однородности выборки коренных зубов корнезубых полёвок, основанный на применении коэффициента вариации, имеет разные количественные рамки для разных стадий гипсодонтности. Так у мезодонтных форм с НН-индексом около 2 – 3,5 заметно повышена изменчивость лингвального тракта задней призмы, что увеличивает изменчивость и всего НН-индекса. В общем случае значения коэффициента вариации НН-индекса от 5 до 15 могут указывать на достаточно однородные выборки. Значения 20 и более указывают на тафономически и/или таксономически смешанные выборки. На рисунке 3 видно, что из общей картины резко выпадают выборки Котловины 2, Нагавской, Ливенцовки 5 и нижних слоев Крыжановки. Это, очевидно, может указывать на значительную тафономическую контаминацию материалов по борзодиям из этих местонахождений. Таким образом, количественная оценка гипсодонтии и анализ её вариабельности позволяют в ряде случаев выявить присутствие переотложенного материала в захоронениях. Микроструктура эмали. Изучена микроструктура эмали группы полевок Borsodia – Prolagurus. Показано закономерное изменение строения эмали зубов, стадии которого могут быть использованы для стратиграфических и эволюционных построений (von Koenigswald, Tesakov, 1997). |
|
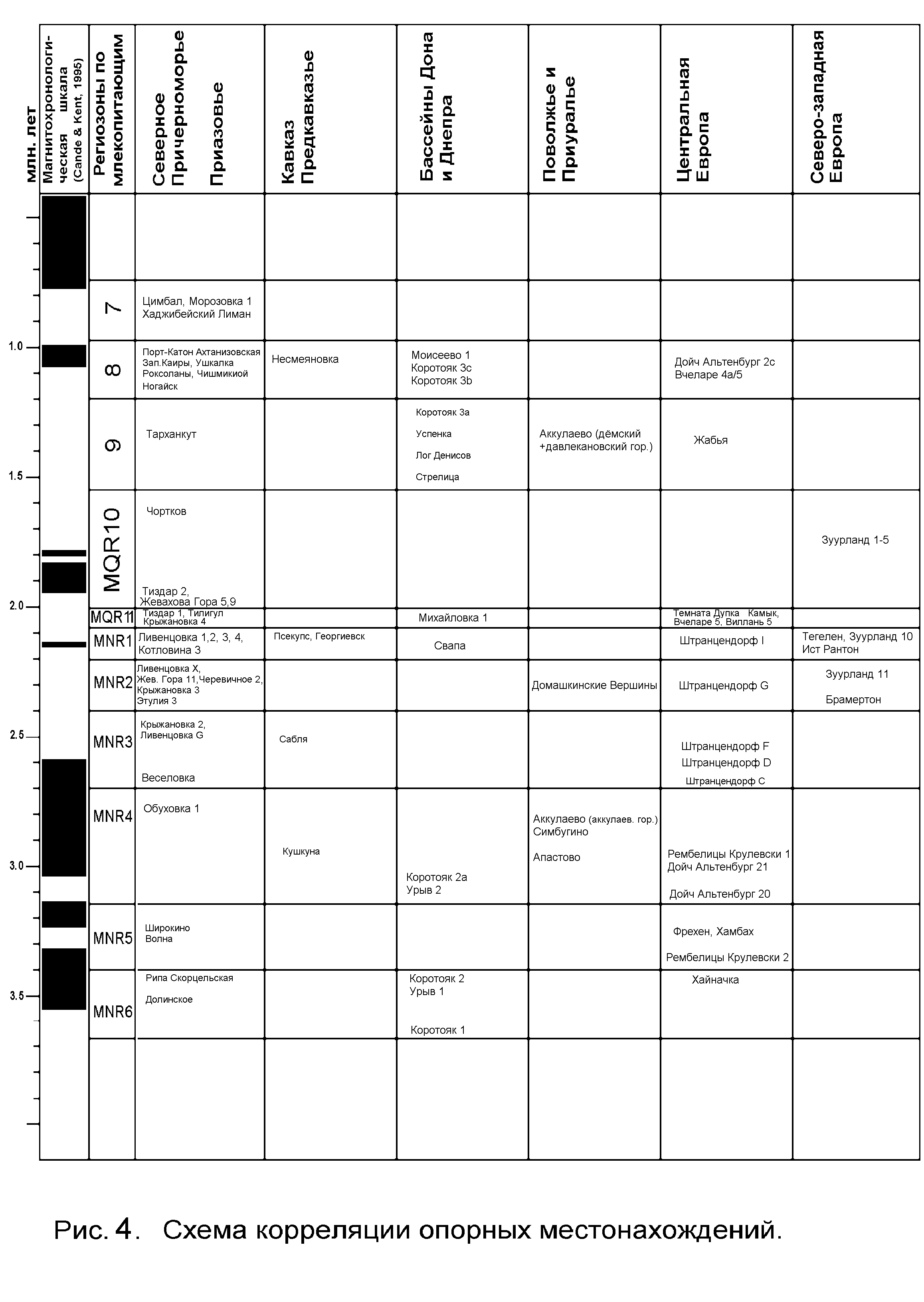
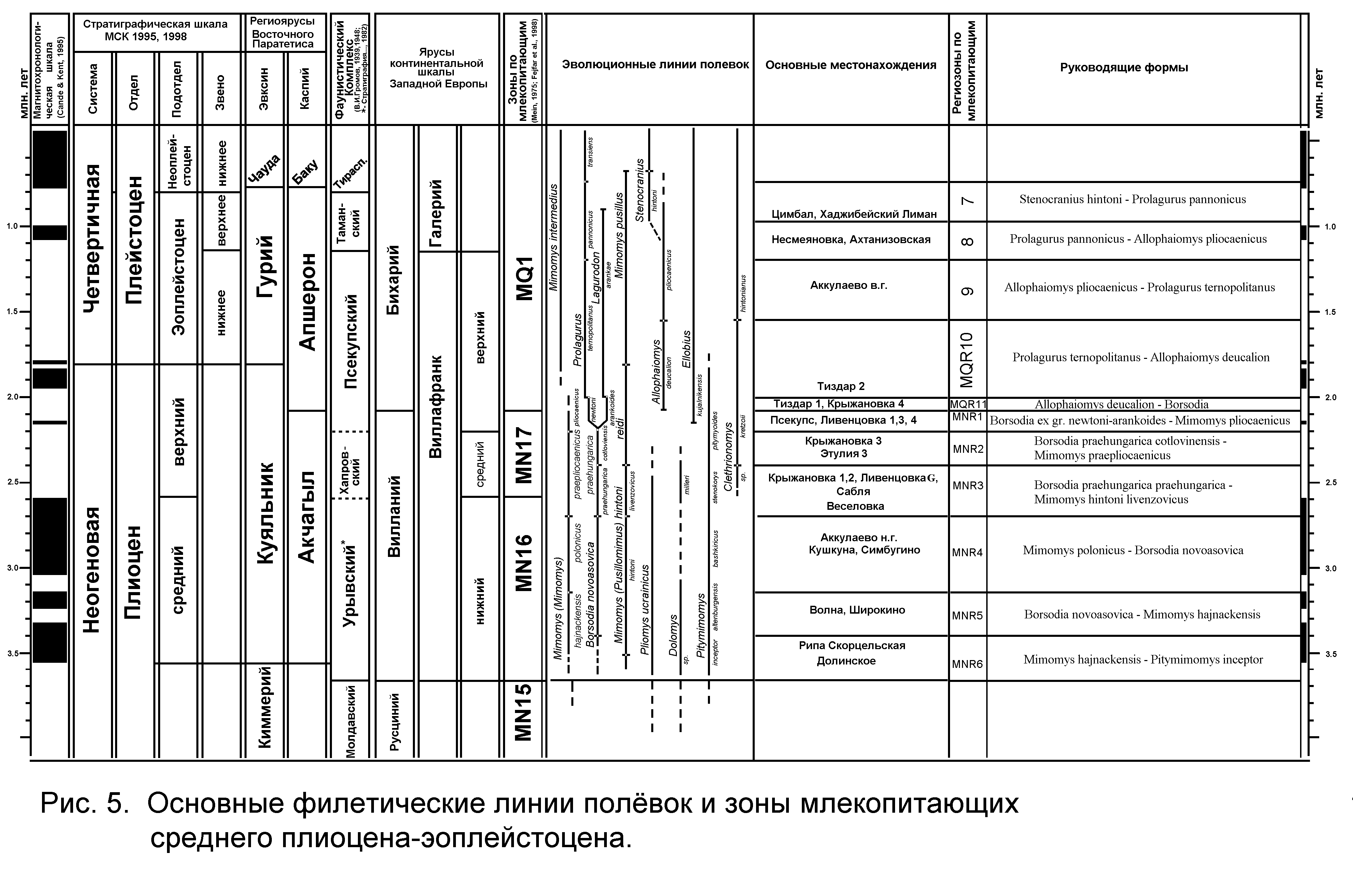
Глава 3. Биостратиграфия среднего плиоцена – эоплейстоцена Восточной Европы на основе анализа эволюционного развития грызунов подсемейства полёвок (Cricetidae, Arvicolinae). 3.1. Последовательность изученных фаун и местонахождений. Для выяснения последовательности фаун и местонахождений и выработки представлений о региональном стратиграфическом подразделении плиоценовых и раннечетвертичных отложений рассмотрены общие особенности ассоциаций мелких млекопитающих, с анализом стадий эволюционного развития форм полёвок (табл. 1). Всего, с учётом многослойных местонахождений, изучено стратиграфическое распространение более 40 таксонов полёвок в 29 фаунистических ассоциациях грызунов из 18 основных и нескольких дополнительных местонахождений. Наиболее примитивные ассоциации мелких млекопитающих представлены в материалах из верхнелевантинских отложений низовьев левобережья Дуная – местонахождения Долинское и Рипа Скорцельская. Их объединяет присутствие примитивных полёвок родов Mimomys, Dolomys, Pliomys. Фауна Рипы Скорцельской по эволюционной продвинутости Dolomys и Pliomys несколько превосходит фауну Долинского. Для фауны Долинского характерен низкокоронковый мелкий вид рода мимомис – Mimomys sp. (НН-индекс менее 1,0), более примитивный, чем Mimomys hintoni Fejfar из Рипы Скорцельской. Линия крупных мимомисов представлена в местонахождении Рипа Скорцельская формой, близкой по эволюционному уровню к типовой M.hajnackensis Fejfar. Фауна этого местонахождения содержит также вид Pitymimomys inceptor sp. nov. (in litt.), более примитивный, чем другие известные виды этой линии, а также первых и наиболее примитивных в изученной последовательности Borsodia cf. steklovi Zazhigin. Следующая группа фаун – Волна, Широкино – отличается присутствием Mimomys hajnackensis Fejfar, более гипсодонтной по сравнению с формой из Рипы Скорцельской. M. hajnackensis из Широкино несколько гипсодонтнее формы из Волны. В Широкино присутствует Mimomys hintoni Fejfar, близкая к типовой форме и более прогрессивная в сравнении с мелкими мимомисами Долинского и Рипы Скорцельской, а также Borsodia novoasovica Top. et Scorik, с НН-индексом от1 до 2 . Далее в рассматриваемой последовательности можно поместить фауны Кушкуны, Симбугина и Аккулаева. Их объединяет присутствие крупной Mimomys polonicus. Форма из Кушкуны, видимо, несколько менее продвинута по сравнению с симбугинской и близка к типовой форме из Рембелиц Крулевских I. В приуральских фаунах представлены также Borsodia novoasovica с НН-индексом от 1,5 до 2,5. Характерны Mimomys hintoni Fejfar и Pitymimomys baschkiricus (Suchov). При этом фауна Симбугина несколько менее продвинута, чем фауна Аккулаева по эволюционному уровню Mimomys hintoni. Следующая группа фаун характеризуется дальнейшим возрастанием гипсодонтии фоновых видов полевок. Сюда можно отнести фауны местонахождений Веселовка, Сабля и Крыжановка 2. Здесь присутствуют Borsodia praehungarica (Schevtschenko) и Mimomys hintoni livenzovicus Alexandrova с НН-индексами от 2,5 до 3,5. В местонахождении Сабля впервые появляется Clethrionomys. Крупный мимомис из этого местонахождения представлен М. praepliocaenicus Rabeder. Отмечены также остатки Pitymimomys ex gr. stenokorys (Rabeder) более продвинутые, чем P. baschkiricus (Suchov) из предшествующей ассоциации. Наиболее молодое местонахождение этой группы, видимо, Крыжановка 2, судя по стадии гипсодонтии Borsodia и мелких мимомисов. Борзодии и мелкие мимомисы, несколько менее развитые по эволюционному уровню, чем формы из Веселовки, присутствуют также в нижних костеносных слоях разрезов Крыжановки и Ливенцовки. Эти формы, возможно, переотложены из более древних отложений. Обратная намагниченность нижних костеносных горизонтов Крыжановки и Ливенцовки, сопоставляемая с началом эпохи Матуяма (Певзнер, 1989, Теса ков, 1993), является дополнительным свидетельством в пользу переотложенности костного материала, поскольку несколько более гипсодонтые формы из разреза Веселовка заключены в прямонамагниченных отложениях (эпоха Гаусс). Ещё более молодой этап выделяется по фаунам Этулии 3, Крыжановки З и Ливенцовки Х. Фауны этого уровня отличаются присутствием продвинутых Borsodia praehungarica cotlovinensis (Top. et Scorik) с НН-индексом от 3,0 до 4,5 и появлением вида Mimomys reidi Hinton с НН-индексом от 3,0 до 4,0. Крупные мимомисы представлены здесь типичными М.praepliocaenicus Rabeder с НН-индексом 4.0 - 4,8. Обычным элементом становятся лесные полевки Clethrionomys kretzoii (Kowalski). Следующая группа фаун представлена в местонахождениях Ливенцовка 4, Псекупс, Георгиевск, Ливенцовка 1-3. Возрастание гипсодонтии мимомисных полевок приводит на этом этапе к появлению очень высококоронковых Borsodia ex gr. newtoni - arankoides со значениями НН-индекса от 4,0 до 4,7. Все эти фауны близки друг к другу также по степени развития мелких мимомисов группы M.reidi (НН-индекс - от 4,0 до 5,0) и лесных полёвок Clethrionomys kretzoii с НН-индексом 2,5 - 3,5. Наименее продвинута в этой группе ассоциация Ливенцовки. Затем идут остальные фауны. В Ливенцовке и Псекупсе присутствует наиболее поздняя форма линии Pitymimomys – P. pitymyoides. Начало нового этапа маркируется появлением некорнезубых полевок Allophaiomys cf. deucalion Kretzoi в недрах мимомисно-борзодиевой ассоциации, еще сходной с отмеченной для предыдущего этапа. Он охарактеризован фаунами местонахождений Тиздар 1 и Крыжановка 4. Массовое появление некорнезубых лагурин рода Prolagurus и их сосуществование с Allophaiomys cf. deucalion отмечает новый фаунистический уровень, представленный фауной Тиздара 2. Последовательность более молодых фаун и местонахождений хорошо контролируется сменяющими друг друга во времени стадиями развития эволюционных линий родов Prolagurus и Allophaiomys-Stenocranius. Выделяется три группы местонахождений (от древних к более молодым): ассоциация Allophaiomys pliocaenicus - Prolagurus ternopolitanus (Аккулаево - верхние горизонты), Allophaiomys pliocaenicus - Prolagurus pannonicus (Ахтанизовская, Несмеяновка), Stenocranius ex gr. hintoni - Prolagurus pannonicus (Цимбал, Хаджибейский лиман). 3.2. Зональные подразделения среднего плиоцена – эоплейстоцена. Рассматриваемый период геологической истории Восточной Европы от примерно 3,5 млн. лет до 0,8 млн. лет назад достаточно подробно охарактеризован фаунами мелких млекопитающих. За более чем 30 лет изучения региона накоплен огромный фактический материал, опубликованы несколько монографических работ по морфологии и систематике, сделаны важные обобщения по истории развития фаун мелких млекопитающих. В нескольких работах предприняты попытки создания биозонального расчленения последовательности фаун и вмещающих отложений и их корреляции с международными схемами и отложениями других регионов (Александрова, 1976, 1989; Топачевский и др. 1987, 1989, 1998; Tesakov, 1993). Для плейстоценового этапа геологической истории Восточной Европы ранее (Вангенгейм, Певзнер, Тесаков, 1998) были выделены региозоны распространения и совместного распространения видов полевок нескольких филетических линий. В настоящей работе предпринята попытка продолжить зональное расчленение на тех же принципах для неогеновой части шкалы. Эволюционным контролем последовательности служат стадии гипсодонтии, взаимопроверяемые по четырем филетическим линиям: Borsodia – Prolagurus, Mimomys ex gr. hintoni – pusillus, Mimomys ex gr. hajnackensis – pliocaenicus и Pitymimomys ex gr. inceptor - pitymyoides. Насколько возможно, были сняты искажения общей картины из-за переотложения материала в некоторых местонахождениях. Для построения шкалы были использованы фауны как изученные автором, так и ряд наиболее важных в стратиграфическом отношении из других районов Восточной Европы. Кроме того, привлекались некоторые опорные местонахождения Центральной и Северо-Западной Европы, которые удалось скоррелировать с восточноевропейскими по близким стадиям эволюционного развития общих форм (Рис. 4). Что касается плейстоценового этапа, то основной целью в данной работе было определить место изученных местонахождений в зональной шкале Восточной Европы, подробно обоснованной в работе автора совместно с Э.А. Вангенгейм и М.А. Певзнером (2001). Плиоценовые зоны обозначаются MNR (млекопитающие неогена России) и нумеруются сверху вниз (1 - 6) как и плейстоценовые зоны – MQR (млекопитающие квартера России) (1 - 11). Ниже рассматривается характеристика зон от более древних к более молодым (Рис. 5). Для определения возраста границ зон использовались главным образом палеомагнитные данные, а также учитывались сведения о малакофауне для местонахождений, приуроченных к морским отложениям. 3.2.1. Плиоценовый этап. Зона MNR6 выделяется как зона совместного распространения Mimomys hajnackensis и Pitymimomys inceptor sp. nov. Критерии границ. Нижняя граница определяется по появлению индекс-видов; верхняя граница - по появлению Borsodia novoasovica. Типовая фауна. Рипа Скорцельская Местонахождения. Рипа Скорцельская, Долинское, Коротояк 1, 2, Урыв 1. Массовый переотложенный материал грызунов этой зоны содержится в среднем костеносном слое местонахождения Котловина (Украина). В Центральной Европе к этой зоне относится местонахождение Хайначка (Словакия) – типовое местонахождение Mimomys hajnackensis. Фаунистическая характеристика. Основным отличием этих фаун от предшествующих русцинийских является мощная диверсификация полевок, в частности появление рода Mimomys (2 филетические линии) с наружным цементом на коренных зубах, родов Borsodia, Villanyia, Pitymimomys, Ungaromys. Часть филетических линий (Dolomys ex gr. nehringi- milleri и Pliomys ex gr. jalpugensis - ucrainicus) продолжает свое развитие, начатое в раннем плиоцене. Все формы полевок имеют очень брахиодонтые коренные, по уровню эволюционного развития близкие к раннеплиоценовым видам «промимомисной» группы конца русциния. Возраст нижней границы несколько древнее инверсии Гильберт – Гаусс (т.е., древнее 3,6 млн. лет). Для определения возраста верхней границы зоны данных пока недостаточно. Корреляция с другими стратиграфическими шкалами показана на Рис. 5. Зона MNR5. Зона совместного распространения Borsodia novoasovica и Mimomys hajnackensis. Критерии границ. Нижняя граница – по появлению Borsodia novoasovica, верхняя – по филетической смене Mimomys hajnackensis на M. polonicus. Типовая фауна. Широкино Местонахождения. Широкино, Волна; в Центральной Европе – Рамбелице Крулевски 2, Фрехен, Хамбах. Фаунистическая характеристика. Доминируют крупные Mimomys hajnackensis со средним значением НН-индекса 2 – 2,5, присутствуют Borsodia novoasovica и Mimomys hintoni. В этой зоне появляется Pitymimomys altenburgensis, продолжается развитие Dolomys и Pliomys. Верхняя граница зоны близка основанию эпизода Каена палеомагнитной эпохи Гаусс. Зона MNR4. Зона совместного распространения Mimomys polonicus и Borsodia novoasovica. Критерии границ. Нижняя граница – по появлению Mimomys polonicus, верхняя – по смене Borsodia novoasovica на Borsodia praehungarica. Типовая фауна. Симбугино. Местонахождения. Кушкуна, Симбугино, Аккулаево (аккулаевский горизонт), Обуховка 1, Апастово, Урыв 2, Коротояк 2а; в Центральной Европе – Дойч Альтенбург 20, 21 – типовое местонахождение для Pitymimomys altenburgensis и Ремблице Крулевски 1 – типовая для Mimomys polonicus. Фаунистическая характеристика. Характерна Mimomys polonicus с НН-индексом от 2,5 до 3,5; мелкие мимомисы представлены M. hintoni с НН-индексом до 2,7, Borsodia novoasovica имеет НН-индекс от 2 до 2,5. В середине зоны Pitymimomys altenburgensis сменяется P. baschkiricus. Возраст верхней границы точно не установлен. Условно она помещается в конце эпохи Гаусс. Зона MNR3. Зона совместного распространения Borsodia praehungarica praehungarica и Mimomys hintoni livenzovicus. Критерии границ. Нижняя граница – по появлению индекс-видов, верхняя – по смене Borsodia praehungarica praehungarica на B. p. cotlovinensis и по смене Mimomys hintoni на Mimomys reidi. Типовая фауна. Веселовка. Местонахождения. Веселовка, Сабля, Крыжановка 2. В местонахождениях Крыжановка 1, Ливенцовка G и Жевахова гора 15 типичная ассоциация грызунов этой зоны найдена в сочетании с переотложенными остатками полёвок предыдущей зоны. В Центральной Европе к этой зоне отнесены местонахождения Штранцендорф C, D, F. Фаунистическая характеристка. Доминируют Borsodia или ассоциация Mimomys – Pitymimomys. Borsodia имеют НН-индекс от 2,5 до 3,5. Mimomys hintoni livenzovicus – от 2,7 до 3,2. Крупные мимомисы представлены M. praepliocaenicus. Впервые появляются Clethrionomys. Pitymimomys представлены P.stranzendorfensis, во второй половине зоны замещающийся P. stenokorys. Присутствуют развитые Dolomys milleri и Pliomys ucrainicus. Возраст верхней границы точно не установлен. Она проходит в ранней части эпохи Матуяма. Зона MNR2. Зона совместного распространения Borsodia praehungarica cotlovinensis и Mimomys praepliocaenicus. Критерии границ. Нижняя граница – по смене Borsodia p. praehungarica на Borsodia praehungarica cotlovinensis и Mimomys hintoni на M. reidi; верхняя – по смене Borsodia praehungarica на B. ex gr. newtoni-arankoides и Mimomys praepliocaenicus на M. pliocaenicus. Типовая фауна. Крыжановка 3 Местонахождения. Крыжановка 3, Этулия 3, Ливенцовка Х, Жевахова гора 11, Черевичное 2, Домашкинские Вершины; в Центральной Европе – Штранцендорф G, в Северо-Западной Европе – Зуурланд 11 и Брамертон. Фаунистическая характеристка. Доминируют Borsodia. В начале зоны исчезают Dolomys и Pliomys ucrainicus. Возрастает разнообразие мимомисной группы: кроме M. praepliocaenicus (НН-индекс от 4 до 4,5), M. reidi (НН-индекс от 3,2 до 4) появляются гипсодонтные мелкие многоцементные формы группы M. tigliensis-tornensis. Присутствуют Pitymimomys stenokorys. Возраст верхней границы предположительно может быть оценен в 2,2 – 2,3 млн. лет. Зона MNR1. Зона совместного распространения Borsodia ex gr. newtoni-arankoides и Mimomys pliocaenicus. Критерии границ. Нижняя граница – по смене Borsodia praehungarica на B. ex gr. newtoni-arankoides и M. praepliocaenicus на M. pliocaenicus. Верхняя граница – по появлению Allophaiomys. Типовая фауна. Псекупс. Местонахождения.Псекупс, Ливенцовка 1 - 4, Георгиевск, Свапа, Котловина 3. В Центральной Европе – Штранцендорф I, в Северо-Западной Европе – Тегелен, Зуурланд 10, Ист Рантон. Фаунистическая характеристика. Особенностью зоны является обособление двух групп борзодий – B. newtoni и B. arankoides (НН-индекс от 4 до 4,7). Характерно разнообразие полёвок мимомисной группы: M. pliocaenicus (НН-индекс 4,5 – 5), M. ex gr. reidi (НН-индекс 4 – 4,6), M. tigliensis, Pitymimomys pitymyoides, многочисленны Clethrionomys kretzoii. Появляются Ellobius (Ellobius), в Центральной и Южной Европе продолжается развитие эндемичных Ungaromys и Pliomys ex gr. episcopalis. Возраст верхней границы твердо не установлен. Она может быть помещена в магнитохронологической шкале между верхней границей эпизода Реюньон II и нижней границей эпизода Олдувей. 3.2.2. Плейстоценовый этап. Кардинальным отличием плейстоценовой фауны мелких млекопитающих от плиоценовой является появление и широкое развитие некорнезубых полёвок, доминирующих в современных фаунах. И хотя зона MQR11 с первыми Allophaiomys (следующая во времени за зоной MNR1) формально относится ещё к плиоцену, представляется целесообразным именно с этой зоны начинать плейстоценовый этап в истории фауны. Зона MQR11. Зона совместного распространения Allophaiomys deucalion и Borsodia. Критерии границ. Нижняя граница – по появлению рода Allophaiomys, верхняя – по появлению рода Prolagurus. Типовая фауна. Крыжановка 4. Местонахождения. Тиздар 1, Крыжановка 4, Тилигул, Михайловка 1, в Центральной Европе – Темната Дупка, Камык, Вилань 5, Вчеларе 5. Фаунистическая характеристика. Зона маркируется в Причерноморье миграционным появлением первых некорнезубых Allophaiomys ex gr. deucalion на фоне существования почти в неизмененном составе комплекса мимомисных полёвок зоны MNR1, в том числе многочисленных поздних Borsodia (НН-индекс 5,5 – 6), доминирующих в этих сообществах. НН-индекс мелких Mimomys превышает 4,7. Верхняя граница зоны древнее эпизода Олдувей и моложе эпизода Реюньон II. Зона MQR10. Зона совместного распространения Prolagurus ternopolitanus и Allophaiomys deucalion. Критерии границ. Нижняя граница – по появлению рода Prolagurus, верхняя – по филетической смене Allophaiomys deucalion на Allophaiomys pliocaenicus. Типовая фауна. Жевахова гора 5,9. Местонохождения . Чортков, Тиздар 2, в Северо-Западной Европе – Зуурланд 1 – 5. Фаунистическая характеристика. Массовые некорнезубые Lagurodon arankae, Prolagurus ternopolitanus и Allophaiomys ex gr. deucalion. Среди мимомисов доминируют Mimomys ex gr. reidi – pusillus. В целом быстро сокращается разнообразие мимомисной группы полёвок. Возраст верхней границы точно не установлен (несколько моложе эпизода Олдувей). Зона MQR9. Зона совместного распространения Allophaiomys pliocaenicus и Prolagurus ternopolitanus. Критерии границ. Нижняя граница – по филетической смене Allophaiomys deucalion на Allophaiomys pliocaenicus и Clethrionomys kretzoii на C. hintonianus, верхняя – по эволюционному замещению Prolagurus ternopolitanus на P. pannonicus. Типовая фауна. Успенка. Местонахождения. Успенка, Лог Денисов, Стрелица 1, Коротояк 3а, Тарханкут, Аккулаево (дёмский и давлекановский горизонты), в Центральной Европе – Жабья пещера. Фаунистическая характеристика. Наряду с массовыми индекс-видами достигают высокой численности Mimomys intermedius и M. pusillus. Обычны Clethrionomys ex gr. hintonianus. В степных районах появляются Eolagurus argyropuloi. Возраст верхней границы примерно оценивается в 1,2 млн. лет (древнее эпизода Харамильо). Зона MQR8. Зона совместного распространения Prolagurus pannonicus и Allophaiomys pliocaenicus. Критерии границ. Нижняя граница – по филетической смене Prolagurus ternopolitanus на Prolagurus pannonicus, верхняя – по появлению Stenocranius hintoni. Типовая фауна. Ногайск. Местонахождения. Ногайск, Коротояк 3b,c, Моисеево 1, Порт-Катон, Западные Каиры, Ушкалка, Роксоланы, Несмеяновка, Ахтанизовская, Чишмикиой, в Центральной Европе – Дойч Альтенбург 2с, Вчеларе 4а/5. Фаунистическая характеристика. Доминируют прогрессивные полиморфные Allophaiomys ex gr. pliocaenicus, Prolagurus pannonicus и P. arankae. Продолжают развитие все остальные филетические линии полёвок, отмеченные для предыдущей зоны. Верхняя граница зоны несколько моложе эпизода Харамильо. Зона MQR7. Зона совместного распространения Stenocranius hintoni и Prolagurus pannonicus. Критерии границ. Нижняя граница – по появлению Stenocranius hintoni, верхняя – по филетической смене Prolagurus pannonicus на Lagurus transiens. Типовая фауна. Карай Дубина. Местонахождения. Карай Дубина, Морозовка 1, Петропавловка, Шамин, Урыв 3а. Фаунистическая характеристика. Массовое появление «микротусных» дериватов Allophaiomys, сокращается численность Mimomys pusillus. В зоне выделяется две подзоны: нижняя (MQR7B) – подзона совместного распространения Stenocranius hintoni и Prolagurus arankae и верхняя (MQR7A) – подзона совместного распространения Pallasiinus protoeconomus и Prolagurus pannonicus. Граница между подзонами проводится по исчезновению Prolagurus (Lagurodon) arankae. Она соответствует границе между стадиями 22 и 21 изотопно-кислородной шкалы (около 865 тыс. лет). Местонахождения. Хаджибейский лиман и Цимбал, изученные автором, относятся к подзоне MQR7B по присутствию Prolagurus arankae. К этой же подзоне принадлежит Морозовка 1. 3.3.Замечания о возрасте ряда фаунистических ассоциаций в некоторых стратиграфических схемах Проведенные исследования фаунистических ассоциаций мелких млекопитающих из опорных местонахождений среднего плиоцена – эоплейстоцена позволили поставить вопрос о необходимости внесения изменений в некоторые стратиграфические схемы. 3.3.1. О горизонтах среднего плиоцена - эоплейстоцена Восточной Европы Для конца среднего плиоцена, верхнего плиоцена и эоплейстоцена К.В.Никифорова с соавторами (1982, 1991 и др.) выделила ряд стратиграфических горизонтов с указанием опорных местонахождений млекопитающих, послуживших для их палеонтологического обоснования: в верхах среднего плиоцена – веселовский (местонахождение Веселовка), в верхнем плиоцене – чистопольский (Симбугино), аккулаевский (Аккулаево), крыжановский (нижние горизонты Крыжановки – 1-3), ферладанский (Ливенцовка 1-4), в эоплейстоцене – домашкинский (Псекупс), бошерницкий (Бошерница), Жеваховский (Жевахова Гора 5,9, Крыжановка 4), ногайский (Приморское ) и морозовский (Морозовка). Фауна большинства из указанных местонахождений была ревизована и проанализирована. Если считать, что эти фауны действительно отражают биостратиграфическую характеристику выделенных горизонтов, то их стратиграфическое положение и последовательность представляются в несколько ином виде (снизу вверх): средний плиоцен – чистопольский, аккулаевский, веселовский; верхний плиоцен - крыжановский, домашкинский, жеваховский; верхний эоплейстоцен – ногайский и морозовский. Бошерницкий горизонт по его фаунистической характеристике может занять любое положение между ногайским и верхней половиной жеваховского горизонта, а ферладанский соответствует домашкинскому горизонту. 3.3.2. Некоторые замечания к биозональной микротериологической схеме В.А.Топачевского с соавторами В.А.Топачевский и его соавторы (1987, 1998 и др.) в течение многих лет развивали и подробно обосновали принцип подразделения последовательности фаун на этапы – ассоциации. В схемах этих авторов в качестве эволюционного контроля последовательности используется степень развития корней по полевкам рода Borsodia (= Villanyia в работах Топачевского и др.). Однако, как указывалось (Гл.2), оценка продвинутости по степени развития корней является недостаточно точной, а главное – не учитывается массовое переотложение ископаемых остатков. Так например, в местонахождении Котловина (средний слой) содержится материал, переотложенный как минимум с двух стратиграфических уровней (см. рис. 1). В местонахождениях Жевахова Гора 15, Крыжановка и Ливенцовка (нижние слои ) наряду с остатками in situ присутствуют формы более древнего стратиграфического уровня. В связи с этим некоторые местонахождения занимают в схеме инвертированное друг относительно друга положение, а характеристика и названия ряда ассоциаций основаны на переотложенных фаунистических элементах. В настоящей работе приведена корреляция последовательности местонахождений схемы В.А.Топачевского и полученная по результатам исследований автора. 3.3.3. О стратиграфическом положении ройвера Нидерландов и Северо-Западной Германии В стратиграфической шкале Северо-Западной Европы к среднему плиоцену относится ройвер. Его нижняя граница в магнитохронологической шкале проводится внутри эпохи Гаусс, а верхняя – в самом начале эпохи Матуяма. Ройвер подразделяется на три части – А, В и С, причем два нижних подразделения имеют нормальную намагниченность, а ройвер С – обратную. В верхней части ройвера B в местонахождениях Хамбах и Фрехен (Германия) собрана богатая фауна мелких млекопитающих с Mimomys hajnackensis. Этот вид характеризует две нижние зоны, выделенные в данной работе для среднего плиоцена (MNR 6 и 5). Граница между зоной MNR5 и более молодой – MNR4 с Mimomys polonicus проводится вблизи основания эпизода Каена эпохи Гаусс. В указанных местонахождениях M. hajnackensis представлена наиболее прогрессивной формой и соответственно должна быть отнесена к концу зоны MNR5. Следовательно, нормально намагниченные отложения верхней части ройвера В могут соответствовать интервалу прямой полярности между эпизодами Мэммос и Каена эпохи Гаусс, а намагниченные обратно отложения ройвера С – эпизоду Каена. Таким образом, верхняя граница ройвера значительно удревняется по сравнению с существующими представлениями. 3.4. Общая характеристика фаун мелких млекопитающих среднего плиоцена - эоплейстоцена Развитие фаун мелких млекопитающих рассматриваемого интервала времени происходило в условиях возрастающей аридизации климата, что отразилось как на изменениях таксономического состава ассоциаций, так и на особенностях эволюционного развития отдельных филетических линий, в частности, на прогрессирующей гипсодонтности полевок, Происходило постепенное увеличение «ксерофильной составляющей» в экологических группировках сначала – в результате увеличения численности немногих видов, а затем – увеличения таксономического разнообразия группы. Анализ таксономического состава и экологических группировок изученных фаун позволяет выделить три крупных этапа в истории фауны мелких млекопитающих среднего плиоцена – эоплейстоцена, совпадающих с подотделами плиоцена и эоплейстоценом. Фауны среднего плиоцена бидоминантны: в Северном Причерноморье – это две филетические линии Mimomys, со-доминант – Borsodia; севернее – доминирование родов Mimomys и Pitymimomys. Фауны верхнего плиоцена остаются бидоминантными, но Borsodia и Mimomys меняются местами – Borsodia становится доминантом. В конце плиоцена важным событием явилось миграционное появление некорнезубой полевки Allophaiomys и эволюционный переход корнезубой Borsodia в некорнезубую Prolagurus. Фауны эоплейстоцена становятся тридоминантными: со-доминируют Allophaiomys, Prolagurus, Mimomys. Постепенно возрастает доминирование некорнезубых полевок и сокращается видовое разнообразие мимомисной группы и численность ее видов. Четкая зональная дифференциация фаун мелких млекопитающих Восточной Европы характерна для всех трех этапов. |
|
Приложение. (Описание палеонтологического материала).
Отряд Rodentia Bowdich, 1821
Род Mimomys F. Major, 1902 Подрод Mimomys F. Major, 1902: M. ex gr. hajnackensis Fejfar, 1961, M. hajnackensis Fejfar, 1961, M. polonicus Kowalski, 1960, M. praepliocaenicus Rabeder, 1981, M. cf. pliocaenicus F.Mayor, 1902. Подрод Pusillomimus Rabeder, 1981: M. sp., M.hintoni hintoni Fejfar, 1961, M.hintoni livenzovicus Alexandrova, 1976, M.reidi Hinton, 1910, M.pusillus von Mйhely, 1914. Род Pitymimomys Tesakov, 1998: P. inceptor sp.nov., P. baschkiricus (Suchov, 1970), P. cf. stenokorys (Rabeder, 1981), P.pitymyoides (Jбnossy et van der Meulen, 1975). Род Borsodia Jбnossy et van der Meulen, 1975: B. ex gr. steklovi (Zazhigin, 1980), B. novoasovica (Topachevski et Scorik, 1977), B. praehungarica praehungarica (Schevtschenko, 1965), B. praehungarica cotlovinensis (Topachevsky et Scorik, 1977), B. newtoni (F.Major, 1902), B. arankoides (Alexandrova, 1976). Род Allophaiomys Kormos, 1932: Allophaiomys cf. deucalion Kretzoi, 1969, Allophaiomys cf. pliocaenicus Kormos, 1932. Род Prolagurus Kormos,1938: Prolagurus (Prolagurus) ternopolitanus (Topachevsky, 1973), Prolagurus (Prolagurus) pannonicus (Kormos, 1930), Prolagurus (Lagurodon) arankae (Kretzoi, 1954). |
|
1. В результате проведенных исследований разработана детальная биостратиграфическая зональная шкала континентальных отложений среднего плиоцена-эоплейстоцена юга Восточной Европы. В основу зональной шкалы положены эволюционные изменения (с учетом количественных оценок морфологических признаков) в четырех филетических линиях полёвок: Borsodia - Prolagurus, Mimomys hajnackensis - pliocaenicus, Mimomys hintoni – pusillus и Pitymimomys inceptor – pitymyoides. 2. Анализ таксономического состава фауны полёвок и геологии опорных местонахождений Причерноморья, Приазовья, Северного Кавказа, изученных автором, а также ревизия фауны важнейших опорных местонахождений Башкирии, Украины и Молдавии позволили в среднем – верхнем плиоцене выделить 6 (MNR1-6), в эоплейстоцене 5 (MQR7-11) зон совместного распространения таксонов. Возраста границ большинства выделенных зон определены на основании главным образом палеомагнитных данных. 3. Все зональные подразделения скоррелированы с магнитохронологической шкалой, с региоярусами стратиграфической шкалы Восточного Паратетиса, а также с западно-европейскими шкалами континентальных отложений. Восточно-европейские местонахождения и фауны скоррелированы с опорными местонахождениями Центральной Европы и некоторыми фаунами Северо-Западной Европы по идентичным эволюционным стадиям руководящих форм. В результате такой корреляции удалось установить, что многие выделенные зоны протягиваются по крайней мере до западных границ Центральной Европы. 4. Установлено, что одним из наиболее надёжных эволюционных показателей для корнезубых полёвок является степень гипсодонтии, выраженная НН-индексом. Причём, изменения этого показателя, прослеженные для четырех филетических линий, позволяют осуществлять взаимоконтроль в определении последовательности ассоциаций. 5. Оценка гипсодонтии полёвок и анализ ее вариабельности позволили предложить количественные критерии выделения форм, переотложенных из стратиграфически более древних горизонтов в аллювиальных захоронениях. 6. Установлено, что некорнезубые полёвки рода Allophaiomys, по появлению которых до последнего времени проводилась неоген-четвертичная граница в континентальных отложениях, на юге Восточной Европы появились раньше - стратиграфически ниже эпизода Олдувей. 7. Ревизия фаун, послуживших для палеонтологического обоснования стратиграфических подразделений в схемах К.В. Никифоровой и др. (1982 и др.), В.А Топачевского и др. (1998) и В. Загвейна (1998), позволила поставить вопрос о внесении некоторых изменений в эти схемы. 8. В систематической части приведены монографические описания 26 видов и подвидов, принадлежащих 7 родам и подродам. Выделен новый род Pitymimomys, в составе которого описан новый вид, характеризующий наиболее архаичную стадию в развитии данной филетической линии. Задачи дальнейших исследований 1. Прослеживание выделенных стратиграфических подразделений на территории смежных регионов Европы и Азии. 2. Поиск викарирующих форм мелких млекопитающих для распространения разработанной зональной шкалы на территорию Южной Европы. 3. Привлечение к стратиграфическим построениям дополнительных филетических линий мелких млекопитающих для увеличения детальности шкалы. 4. Разработка биозональной шкалы для нижнего плиоцена. 5. Монографическое других описание стратиграфически значимых форм мелких млекопитающих. |
|
1. Тесаков А.С. Магнитостратиграфические и биостратиграфические исследования в страторегионе псекупского фаунистического комплекса. Бюлл. Комис. по изучению четвертичного периода, N 59, 1990, сс. 81-93 (М.А.Певзнер, Э.А. Вангенгейм - соавторы).
2. Тесаков А.С. Положение таманского фаунистического комплекса в
стратиграфической и магнитостратиграфической шкалах. Бюлл. Комис. пр изучению четвертичного периода, 1991, № 60, с. 41-52 (Э.А. Вангенгейм, М.Л. Векуа и др. 3. Tesakov A.S. Evolution of Borsodia (Arvicolidae, Mammalia) in the Villanyian and in the Early Biharian. Quaternary International, 1993, vol.19, pp. 41-45. 4. Тесаков А.С. Позднеплиоценовые и раннеплейстоценовые мелкие млекопитающие юга Восточной Европы. Стратиграфия. Геологическая корреляция. 1993, T.I, № 3, сс. 92-96. 5. Тесаков А.С. Первая находка Mimomys pitymyoides (Mammalia, Rodentia) на Северном Кавказе и ее стратиграфическое значение. Бюлл. комис. изуч. четвертич. периода, 1994, № 61, сс. 43-46. 6. Тесаков А.С. Возрастные соотношения отложений плиоценовых аллювиальных равнин междуречья Прут - Южный Буг. Стратиграфия. Геологическая корреляция, 1995, 3, № 1, сс. 61-72. (Э.А. Вангенгейм, М.А. Певзнер – соавторы). 7. Tesakov A.S. Evolution of small mammal communities from the south of Eastern Europe near the Pliocene-Pleistocene boundary. Acta Zoologica Cracoviensia, 1995, vol. 38, no. 1, pp. 121- 127. 8. Tesakov A.S. Ruscinian from the territory of the former Soviet Union. Newsl. Stratigraphy, 1996, 33 (2), pp. 77-97. (M.A. Pevzner, E.A. Vangengeim, I.A. Vislobokova, M.V. Sotnikova – соавторы). 9. Tesakov A.S. Evolution of bank voles (Clethrionomys, Arvicolinae) in late Pliocene and early Pleistocene of Eastern Europe. Acta Zoologica Cracoviensia, 1996, vol. 39, 1, pp. 541-547. 10. Tesakov A.S. The evolution of the schmelzmuster of Lagurini (Arvicolinae, Rodentia), Palaeontographica B, 1997, 245, pp. 45-61. (von Koenigswald W. – соавтор). 11. Tesakov A.S. Voles of the Tegelen fauna, Mededelingen Nederlands Instituut voor Toegepaste Geowetenschappen TNO, 1998, № 60, рр. 71-134. 12 Tesakov A.S. Evolution of Plio-Pleistocene voles with the special reference to demographic features of fossil assemblages. Mededelingen Nederlands Instituut voor Toegepaste Geowetenschappen TNO, 1998, № 60, рр. 555-564. (Kazantseva N.E. - соавтор). 13. Tesakov A.S. The early Pleistocene Mimomys hordijki sp. nov. from the Zuurland borehole (The Netherlands) Mededelingen Nederlands Instituut voor Toegepaste Geowetenschappen TNO, 1998, № 63, pp. 187-192 (van Kolfschoten T. – соавтор). 14. Tesakov A.S. Introductory remarks: Does Allophaiomys exist? Paludicola, 1998, vol. 2, no.l, pp. 1-7. (MartinR.A – соавтор). 15. Tesakov A.S. Early stage of Allophaiomys evolution in eastern Europe. Paludicola, 1998, vol. 2, no. l, pp. 98-105. 16. Tesakov A.S. The position of the Tizdar Locality (Taman Peninsula, Russia) in the magnetochronological scale. Paludicola, 1998, vol. 2, no. 1, pp. 95-97. (Pevzner M.A., Vangengem E.A. – соавторы).
17. Тесаков А.С. Находки древнейших некорнезубых полёвок родов Allophaiomys и
Prolagurus на территории Восточной Европы. Докл. РАН, 1999, т. 366, № 4, сс. 452-455 18. Тесаков А.С. Некоторые замечания по фауне мелких млекопитающих Домашкинских Вершин (сборы Л.П.Александровой 1973 г.) - В кн.: Яхимович В.Л., А.Г.Яковлев (ред.): Опорный разрез плиоцена и плейстоцена Домашкинские Вершины. Уфа, 2000, сc. 70-71.
19. Тесаков А.С. Зональное расчленение квартера Восточной Европы по мелким
млекопитающим. Стратиграфия. Геологическая корреляция. 2001, т.9, №3, сс. 76-88 20. Tesakov A.S. New data on Plio-Pleistocene jerboas of the genus Plioscirtopoda from Eastern Europe (Mammalia, Dipodidae). Zoosystematica Rossica, 2001, v.9, pp. 237-244. 21. Tesakov A.S. Quaternary zonal subdivision of Eastern Europe based on vole evolution. Boll. Soc. Paleontol. Italiana, 2001, v.40, n. 2, pp. 269-274 (Pevzner M. A.,Vangengeim E.A.- coавторы). 22. Tesakov A.S. The sequence of Plio-Pleistocene mammal faunas from the south Russian Plain (the Azov Region). Boll. Soc. Paleontol. Italiana, 2001, v.40, n. 2, p.133-138 (Baigusheva V.S., Titov V.V.- соавторы). Работы, сданные в печать: 23. Тесаков А.С. Положение Mimomys baschkirica Suchov, 1970 в системе мимомисных полевок. Сборник ЗИН РАН. 24. Тесаков А.С. Расчленение плейстоцена по фауне млекопитающих. Труды Всероссийского совещания по изучению четвертичного периода, ВСЕГЕИ (Вангенгейм Э.А. и Певзнер М.А. – соавторы). 25. Тесаков А.С. Время появления первых некорнезубых полёвок родов Allophaiomys и Prolagurus. Труды Всероссийского совещания по изучению четвертичного периода, ВСЕГЕИ (Вангенгейм Э.А. и Певзнер М.А. – соавторы). 26. Тесаков А.С. О горизонтах верхнего плиоцена и эоплейстоцена Восточной Европы. Труды Всероссийского совещания по изучению четвертичного периода, ВСЕГЕИ (Вангенгейм Э.А. и Певзнер М.А. – соавторы). 27. Tesakov A. Position of reference mammal localities of the Lower Villafranchian in the magnetochronological time scale. Lynx. (Vangengeim E., Pevzner M. – соавторы). |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}